Переход от бодрствования ко сну предполагает два возможных пути. Прежде всего не исключено, что механизмы, поддерживающие состояние бодрствования, постепенно «утомляются». В соответствии с такой точкой зрения сон - это пассивное явление, следствие снижения уровня бодрствования. Однако не исключено и активное торможение механизмов, обеспечивающих бодрствование. И.П. Павлов выделял два механизма развития сна, которые, по существу, подтверждают правомерность позиций сторонников как пассивной, так и активной теории сна. С одной стороны, сон возникает как явления охранительного торможения в результате сильного и длительного раздражения какого-либо отдельного участка коры больших полушарий. С другой стороны, сон возникает как результат внутреннего торможения, т.е. активного процесса формирования отрицательного условного рефлекса. Важную роль в регуляции цикла сон – бодрствование играет ретикулярная формация ствола мозга, где находится множество диффузно расположенных нейронов, аксоны которых идут почти ко всем областям головного мозга, за исключением неокортекса. Роль РФ в цикле сон – бодрствование была исследована в конце 1940-х годов учеными Г. Моруцци и Н. Мэгуном, обнаружившими, что высокочастотное электрическое раздражение этой структуры у спящих кошек приводит к их мгновенному пробуждению. И напротив, повреждения ретикулярной формации вызывают постоянный сон, напоминающий кому; перерезка же только сенсорных трактов, проходящих через ствол мозга, такого эффекта не дает. Наиболее ранними теориями сна были гуморальные. Фактор сна, лишенный видовой специфичности, был выделен из ликвора коз, подвергнувшихся депривации сна. Согласно сосудистой (циркуляторной или гемодинамической) теории сна, наступление сна связано со снижением кровотока в мозге или с его усилением. Современные исследования показали, что в течение сна действительно происходит колебание кровенаполнения мозга. Р. Лежандр и X. Пьерон (1910) считали, что сон возникает в результате накопления токсических продуктов обмена вследствие утомления (гипотоксины). Собакам долгое время не давали спать, а затем забивали, экстрагировали вещества из мозга и вводили другим собакам. У последних развивались признаки крайнего утомления и возникал глубокий сон. То же наблюдалось при «переносе» сыворотки крови или спинно-мозговой жидкости.
В 1965 г. Ж. Монье использовал препарат с перекрестным кровообращением у двух кроликов, т.е. кровь от мозга одного кролика попадала в туловище другого. Если у одного из кроликов раздражали участки мозга, вызывающие сон, то второй кролик тоже засыпал. Во время сна в мозгу обнаружено избыточное накопление ряда биологически активных веществ – ацетилхолина, гаммааминомасляной кислоты, серотонина.
В середине XIX в. большее распространение получили нервные теории сна. Один путь исследований – это изучение так называемого центра сна, о существовании которого в ядрах гипоталамуса у животных утверждал швейцарский физиолог В. Гесс (1933). У больных с пораженным гипоталамусом также отмечалась повышенная сонливость: у раненого солдата осколок снаряда находился на уровне гипоталамуса, а попытка извлечь осколок пинцетом вызывала мгновенный глубокий сон.
Выделяют так называемую информационную теорию сна (Н. Вингер). Согласно этой теории, в течение дня мозг накапливает огромную информацию, усвоение которой затруднено, а часть ее не имеет отношения к долговременным задачам. Если кратковременная память заполняется днем, то ночью часть содержащейся в ней информации переписывается в долговременную память. Особенности процессов переработки информации требуют отключения от сигналов внешнего мира.
В верхних отделах ствола мозга есть две области – ядра шва и голубое пятно, у нейронов которых такие же обширные проекции, как и у нейронов ретикулярной формации, т.е. достигающие многих областей ЦНС. Ядра шва захватывают срединную часть продолговатого мозга, моста и среднего мозга. Разрушение их устраняет синхронизацию ЭЭГ и медленный сон. С помощью специальной методики флуоресценции гистохимики показали, что нейроны ядер шва синтезируют серотонин и направляют его через свои аксоны к ретикулярной формации, гипоталамусу, лимбической системе. Серотонин – тормозной медиатор моноаминергической системы мозга. Блокада синтеза серотонина устраняет у кошки медленный сон, у которой сохраняется лишь парадоксальный сон.
В среднем мозге (покрышка) обнаружено скопление нейронов, синтезирующих норадреналин (голубое пятно). Стимуляция голубого пятна вызывает торможение нейронной активности во многих структурах мозга при росте двигательного возбуждения животного и ЭЭГ-десинхронизации. Полагают, что активирующее влияние голубого пятна осуществляется через механизм торможения тормозных интернейронов. Ядра шва и голубое пятно действуют как антагонисты. Медиатором в клетках ядер шва служит серотонин (5-гидрокситриптамин, 5-НТ), а голубого пятна – норадреналин. Разрушение ядер шва у кошки приводит к полной бессоннице в течение нескольких дней; но за несколько последующих недель сон нормализуется. Частичная бессонница может быть также вызвана подавлением синтеза 5-НТ п -хлорфенилаланином. Ее можно устранить введением 5-гидрокситриптофана, предшественника серотонина (последний не проникает через гематоэн-цефалический барьер). Двустороннее разрушение голубого пятна приводит к полному исчезновению БДГ-фаз, не влияя на медленноволновый сон. Истощение запасов серотонина и норадреналина под влиянием резерпина вызывает, как и следовало ожидать, бессонницу. Однако оказалось, что нейроны ядер шва наиболее активны и выделяют максимум серотонина не во время сна, а при бодрствовании. Кроме того, возникновение БДГ, по-видимому, обусловлено активностью нейронов не столько голубого пятна, сколько более диффузного подголубого ядра. Судя по результатам недавних экспериментов, серотонин служит и медиатором в процессе пробуждения, и «гормоном сна» в бодрствующем состоянии, стимулируя синтез или высвобождение «веществ сна» (факторов сна), которые в свою очередь вызывают сон. Структуры таламуса выполняют функцию «пейсмекера» для вызова ритмических потенциалов веретен во сне и a-ритма в бодрствовании. Таламокортикальный механизм можно рассматривать как механизм внутреннего торможения, способного изменять активность мозга частично или глобально таким образом, что сенсорные, моторные и высшие функции мозга подавляются.
Рис. 9.4. Реципрокные отношения мультинейронной активности ретикулярной формации и преоптической области гипоталамуса в различных фазах цикла бодрствование–сон; калибровка: 40 мкВ, 50 мс (по Т.Н. Ониани, 1983, с изменениями)
Структуры, ответственные за медленный сон, находятся в каудальной части мозгового ствола, главным образом – в продолговатом мозге. Наличие сходных гипногенных структур было установлено также и в задней части моста. Двигательные и ЭЭГ-проявления фазы парадоксального сна связаны с активацией структур в области моста. Эта фаза сна сокращается при эмоциональном стрессе, при этом удлиняется период засыпания.
Рядом с голубым пятном имеется группа гигантских ретикулярных нейронов, которые направляют свои аксоны вверх и вниз к различным структурам мозга. В бодрствовании и медленном сне эти нейроны малоактивны, но их активность весьма высока во время парадоксального сна.
Были сделаны попытки обнаружить особые вещества либо после длительного лишения сна, либо у спящего человека. Первый из этих подходов основан на предположении о том, что фактор(ы) сна во время бодрствования накапливаются до вызывающего сон уровня, а второй – на гипотезе, согласно которой они образуются или выделяются во сне.
Оба подхода дали определенные результаты. Так, при проверке первой гипотезы из мочи и спинномозговой жидкости человека и животных был выделен небольшой глюкопептид – фактор S, вызывающий медленноволновый сон при введении другим животным. Существует, по-видимому, и фактор сна с БДГ. Второй подход привел к открытию индуцирующего глубокий сон нонапептида (в настоящее время он уже синтезирован), так называемого пептида D-сна (DSIP , delta-sleep inducing peptide). Однако пока неизвестно, играют ли эти и многие другие «вещества сна», обнаруженные при проверке обеих гипотез, какую-либо роль в его физиологической регуляции. Более того, выделенные пептиды часто вызывают сон лишь у животных определенного вида; кроме того, он возникает и под действием других веществ.
Однако сросшиеся девочки-близнецы могли спать порознь, что свидетельствует о второстепенной роли гуморальных факторов и решающей роли в развитии сна нервной системы.
Развивается представление о том, что цикл бодрствование – сон обеспечивается системой двух центров. К. Экономо на основе клинических наблюдений больных с повреждениями различных участков гипоталамуса предположил, что центр бодрствования локализован в заднем, а центр сна – в его передних отделах. С. Рэнсон, производя локальные повреждения различных участков гипоталамуса, подтвердил это мнение. В настоящее время считают, что гипоталамус является критической зоной для регулирования цикла бодрствование – сон. Это мнение подтверждается и тем, что как высокочастотное, так и низкочастотное электрическое раздражение преоптической области гипоталамуса вызывает синхронизацию электроэнцефалограммы и поведенческий сон. Противоположный эффект, а именно поведенческое и электроэнцефалографическое пробуждение Т.Н. Ониани наблюдал при раздражении заднего гипоталамуса. Это позволяет предположить наличие реципрокного взаимоотношения между передней и задней областями гипоталамуса и его значение для регуляции чередования различных фаз цикла бодрствование – сон. По данным Т.Н. Ониани, в цикле бодрствование – сон мультинейронная активность ретикулярной формации (активирующей системы) и преоптической области гипоталамуса (тормозной системы) меняется реципрокно (рис. 9.4).
Гипноз
Гипноз – состояние человека, вызванное искусственно с помощью внушения, когда не полностью выключено сознание и снижены реакции на все раздражители, кроме слов гипнотизирующего, к которым человек становится чрезвычайно восприимчивым. В ответ на многократные или монотонные стимулы непременно развивается внутреннее торможение. Если такая стимуляция продолжается, то наступает сон. Переходный период между бодрствованием и сном назван гипнотическим состоянием. И.П.Павлов считал, что гипнотическое состояние является частичным сном. Он разделил гипнотическое состояние на три фазы.
Первая фаза называется уравнительной, в это время сильные и слабые стимулы вызывают одинаковые условные ответы.
Парадоксальная фаза характеризуется более глубоким сном, слабые раздражители вызывают более интенсивный ответ, чем сильные.
Ультрапарадоксальная фаза означает еще более глубокий сон, когда ответ вызывают только слабые стимулы, а сильные приводят к еще большему распространению торможения. За этими тремя фазами следует глубокий сон.
Однако оказалось, что в гипнозе волны электроэнцефалограммы (ЭЭГ) сходны с ЭЭГ бодрствования, а не сна. Выяснилось, что одни люди более подвержены гипнозу, чем другие, а некоторые вовсе не поддаются ему (что говорит о различном соотношении процессов возбуждения и торможения у разных людей). В последние годы получили распространение теории, согласно которым гипнотическое состояние нельзя приравнивать ни ко сну, ни к бодрствованию.
Обычно выделяют четыре стадии гипноза.
1. Стадия гипноидности, сопровождающаяся психическим и мышечным расслаблением, миганием и закрыванием глаз.
2. Стадия легкого транса, для которой характерна каталепсия конечностей (конечности могут длительное время находиться в необычном положении).
3. Стадия среднего транса, когда возникает амнезия, изменения личности, возможны простые гипнотические внушения.
4. Стадия глубокого транса, которая характеризуется полным сомнамбулизмом и фантастическими внушениями.
При гипнозе изменяется состояние коры больших полушарий, которое меняет соотношение между первой и второй сигнальными системами. Снижается контроль второй сигнальной системы над первой и облегчается воспроизведение пережитых состояний и зрительных образов. Возбуждение коры при восприятии слов гипнотизера оказывается изолированным от влияния остального мозга, воздействия на него оказываются необычайно сильными с длительным последействием, что обусловливает и длительный лечебный эффект гипнотического внушения (гипнотерапии).
Во время гипноза высшая нервная деятельность существенно меняется. Это касается сенсорной сферы, когда можно внушить отсутствие слуха, зрения, осязания и даже боли, можно получить извращенные реакции (иллюзии), например на вкусовые или обонятельные раздражения, вызвать галлюцинации – восприятие несуществующих предметов и явлений (положительные) или выключить из восприятия определенные предметы (отрицательные галлюцинации). Принципиальный интерес представляют исследования с внушением возраста, когда у человека восстанавливаются особенности памяти, речи, почерка, характерные для его детства, меняется фонетическая выразительность речи, темп и содержание мышления.
Гипнотику можно внушить различные эмоциональные состояния, воздействовать на интеллектуальные, моральные и этические чувства, на сферу его внимания, но нельзя изменить личность человека вопреки его социальным и индивидуальным убеждениям.
Механизмы гипноза остаются неизвестными, а завершенной теории гипноза еще не разработано, хотя ряд заболеваний успешно излечивается под гипнозом.

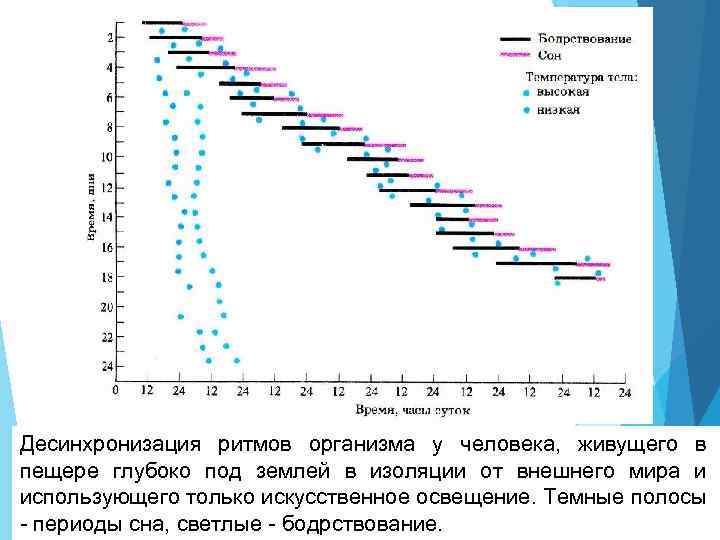 Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира и использующего только искусственное освещение. Темные полосы - периоды сна, светлые - бодрствование.
Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира и использующего только искусственное освещение. Темные полосы - периоды сна, светлые - бодрствование.
 Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
 Бодрствование - состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. В поддержании состояния бодрствования важнейшую роль играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование (обычное состояние не спящего человека) образует поле всевозможных сочетаний функций сознания - от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.
Бодрствование - состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. В поддержании состояния бодрствования важнейшую роль играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование (обычное состояние не спящего человека) образует поле всевозможных сочетаний функций сознания - от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.


 Сон – это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
Сон – это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
 Выделяют две основные стадии сна - «медленный» и «быстрый» сон. Феномен «быстрого» сна открыли в 1953 г. американские ученые А. Азеринский и К. Клейтман. Медленноволновой сон человека периодически прерывается короткими периодами низкоамплитудной ЭЭГ (похожей на десинхронизацию ЭЭГ при пробуждении), сопровождающимися быстрыми движениями глазных яблок. Отсюда часто употребляемое название этой стадии - парадоксальная, или стадия быстрых движений глаз (БДГ-сон, или REM-стадия - от rapid eye movement). Люди, разбуженные во время парадоксальной стадии сна, в 80 % случаев сообщали о сновидениях. Мышцы тела расслаблены сильнее всего во время парадоксального сна, однако вегетативные показатели (дыхание, сердцебиение, кровяное давление и др.) во время парадоксального сна могут соответствовать активному бодрствованию (вплоть до так называемой «вегетативной бури»).
Выделяют две основные стадии сна - «медленный» и «быстрый» сон. Феномен «быстрого» сна открыли в 1953 г. американские ученые А. Азеринский и К. Клейтман. Медленноволновой сон человека периодически прерывается короткими периодами низкоамплитудной ЭЭГ (похожей на десинхронизацию ЭЭГ при пробуждении), сопровождающимися быстрыми движениями глазных яблок. Отсюда часто употребляемое название этой стадии - парадоксальная, или стадия быстрых движений глаз (БДГ-сон, или REM-стадия - от rapid eye movement). Люди, разбуженные во время парадоксальной стадии сна, в 80 % случаев сообщали о сновидениях. Мышцы тела расслаблены сильнее всего во время парадоксального сна, однако вегетативные показатели (дыхание, сердцебиение, кровяное давление и др.) во время парадоксального сна могут соответствовать активному бодрствованию (вплоть до так называемой «вегетативной бури»).

 График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального сна (БДГ) - от 8 часов у новорожденных до 1 часа у пожилых людей. Изменения в продолжительности медленноволнового сна (МВ) не столь выражено; уменьшение от 8 часов до примерно 5 часов.
График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального сна (БДГ) - от 8 часов у новорожденных до 1 часа у пожилых людей. Изменения в продолжительности медленноволнового сна (МВ) не столь выражено; уменьшение от 8 часов до примерно 5 часов.
 ЭЭГ человека при бодрствовании и сне Бодрствование Частота Амплитуда Синхронизация (регулярность колебаний) Медленноволновый сон Парадоксальный сон (БДГ) высокая низкая (нерегулярная активность, десинхронизация) высокая (синхронизированная активность) низкая (нерегулярная активность, десинхронизация)
ЭЭГ человека при бодрствовании и сне Бодрствование Частота Амплитуда Синхронизация (регулярность колебаний) Медленноволновый сон Парадоксальный сон (БДГ) высокая низкая (нерегулярная активность, десинхронизация) высокая (синхронизированная активность) низкая (нерегулярная активность, десинхронизация)
 В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне. В процессе засыпания человек проходит 4 стадии, от спокойного бодрствования с закрытыми глазами (альфа-ритм) до собственно медленноволнового сна (дельта-ритм).
В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне. В процессе засыпания человек проходит 4 стадии, от спокойного бодрствования с закрытыми глазами (альфа-ритм) до собственно медленноволнового сна (дельта-ритм).
 Основные ритмы ЭЭГ человека Частота Состояние 12 -25 Гц Бодрствование с открытыми глазами и БДГ-сон Альфа-ритм 8 -12 Гц Бодрствование с закрытыми глазами Тета-ритм 4 -8 Гц Засыпание 1 -4 Гц Медленноволновый сон Бета-ритм Дельта-ритм
Основные ритмы ЭЭГ человека Частота Состояние 12 -25 Гц Бодрствование с открытыми глазами и БДГ-сон Альфа-ритм 8 -12 Гц Бодрствование с закрытыми глазами Тета-ритм 4 -8 Гц Засыпание 1 -4 Гц Медленноволновый сон Бета-ритм Дельта-ритм
 ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных мышц.
ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных мышц.
 Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки Бремера» - по Бремеру, 1937). При перерезке на границе головного мозга со спинным (препарат А) в ЭЭГ сохранялась картина бодрствования, а при перерезке на уровне среднего мозга (препарат Б) получали препарат спящего мозга. Бремер заключил, что в изолированном головном мозгу имеется центр пробуждения, расположенный в продолговатом и среднем мозгу.
Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки Бремера» - по Бремеру, 1937). При перерезке на границе головного мозга со спинным (препарат А) в ЭЭГ сохранялась картина бодрствования, а при перерезке на уровне среднего мозга (препарат Б) получали препарат спящего мозга. Бремер заключил, что в изолированном головном мозгу имеется центр пробуждения, расположенный в продолговатом и среднем мозгу.
 Дж. Моруцци и Х. Мегун показали в 1949 г. , что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон при этом в то время рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включением» таламокортикальных синхронизирующих процессов.
Дж. Моруцци и Х. Мегун показали в 1949 г. , что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон при этом в то время рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включением» таламокортикальных синхронизирующих процессов.
 В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря работам Бремера, Моруцци, Мегуна и других исследователей), существует большое количество центров, которые различаются по своим функциям, выделяемым нейромедиаторам и локализации в мозге. Большинство из них действительно находятся в ретикулярной формации ствола мозга или вблизи нее, но часть из них – в гипоталамусе и других структурах мозга.
В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря работам Бремера, Моруцци, Мегуна и других исследователей), существует большое количество центров, которые различаются по своим функциям, выделяемым нейромедиаторам и локализации в мозге. Большинство из них действительно находятся в ретикулярной формации ствола мозга или вблизи нее, но часть из них – в гипоталамусе и других структурах мозга.
 Некоторые системы регуляции сна и бодрствования. Видно, как в цепях, выходящих из голубого пятна (медиатор - норадреналин) и ядер шва (медиатор - серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
Некоторые системы регуляции сна и бодрствования. Видно, как в цепях, выходящих из голубого пятна (медиатор - норадреналин) и ядер шва (медиатор - серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
 Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна Бодрствование Медленноволновый сон Парадоксальный сон Преобладающий ритм в ЭЭГ бета-ритм дельта-ритм бета-ритм Выделение норадреналина (из голубого пятна) максимально снижено отсутствует Выделение серотонина (из дорзальных ядер шва) максимально снижено отсутствует Выделение гистамина (из туберомамиллярного ядра заднего гипоталамуса) максимально снижено отсутствует Выделение ацетилхолина (из базальных ядер переднего мозга - базальное крупноклеточное ядро и др.) максимально отсутствует максимально
Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна Бодрствование Медленноволновый сон Парадоксальный сон Преобладающий ритм в ЭЭГ бета-ритм дельта-ритм бета-ритм Выделение норадреналина (из голубого пятна) максимально снижено отсутствует Выделение серотонина (из дорзальных ядер шва) максимально снижено отсутствует Выделение гистамина (из туберомамиллярного ядра заднего гипоталамуса) максимально снижено отсутствует Выделение ацетилхолина (из базальных ядер переднего мозга - базальное крупноклеточное ядро и др.) максимально отсутствует максимально
 В настоящее время очевидно, что бодрствование и сон – качественно различные состояния, каждое из которых имеет свои управляющие центры.
В настоящее время очевидно, что бодрствование и сон – качественно различные состояния, каждое из которых имеет свои управляющие центры.
 Предполагаемые центры бодрствования: 1 - базальные ядра переднего мозга (выделяют ацетилхолин), 2 - ядра заднего гипоталамуса (туберомаммиллярное ядро) (выделяют гистамин), 3 - дорзальные ядра шва (выделяют серотонин), 4 - область покрышки моста (выделяет ацетилхолин и глутамат), 5 - голубое пятно (выделяет норадреналин). Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным). Орексиновые нейроны немногочисленны и расположены в гипоталамусе Предполагается, что именно выделение нейропептида орексина нейронами этих ядер переключает мозг в режим бодрствования.
Предполагаемые центры бодрствования: 1 - базальные ядра переднего мозга (выделяют ацетилхолин), 2 - ядра заднего гипоталамуса (туберомаммиллярное ядро) (выделяют гистамин), 3 - дорзальные ядра шва (выделяют серотонин), 4 - область покрышки моста (выделяет ацетилхолин и глутамат), 5 - голубое пятно (выделяет норадреналин). Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным). Орексиновые нейроны немногочисленны и расположены в гипоталамусе Предполагается, что именно выделение нейропептида орексина нейронами этих ядер переключает мозг в режим бодрствования.
 Функция орексина как медиатора, ответственного за координацию других центров бодрствования, была открыта совсем недавно – около 2003 г. , а сам орексин был открыт лишь в 1998 г. При потере хотя бы части орексиновых нейронов возникает нарколепсия – дневная сонливость.
Функция орексина как медиатора, ответственного за координацию других центров бодрствования, была открыта совсем недавно – около 2003 г. , а сам орексин был открыт лишь в 1998 г. При потере хотя бы части орексиновых нейронов возникает нарколепсия – дневная сонливость.
 Схема центра расположения центра медленного сна. Вверху – бодрствование, когда центр сна заторможен и ретикулярная формация (РФ) активирует кору; внизу – центр сна возбужден, РФ заторможена и кора не активируется.
Схема центра расположения центра медленного сна. Вверху – бодрствование, когда центр сна заторможен и ретикулярная формация (РФ) активирует кору; внизу – центр сна возбужден, РФ заторможена и кора не активируется.
 Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор - гаммааминомасляная кислота (ГАМК); активность нейронов незначительна в бодрствовании, но резко возрастает в период обычного сна и прекращается во время парадоксального. Центр парадоксального сна: ретикулярная формация покрышки моста (латеродорзальная / педункуло-понтийная области покрышки моста), медиаторы - ацетилхолин и глутамат.
Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор - гаммааминомасляная кислота (ГАМК); активность нейронов незначительна в бодрствовании, но резко возрастает в период обычного сна и прекращается во время парадоксального. Центр парадоксального сна: ретикулярная формация покрышки моста (латеродорзальная / педункуло-понтийная области покрышки моста), медиаторы - ацетилхолин и глутамат.
 Список Литературы Дубынин В. А. Регуляторные системы организма человека. М. : Дрофа, 2003. Дудьев В. П. Психомоторика: cловарь-справочник, 2008 г. Мэгун Г. Бодрствующий мозг. М. : Мир, 1965. Росси Я. А. , Цанкетти А. Ретикулярная формация ствола мозга. М. : ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М. : Изд-во Моск. ун-та, 1987.
Список Литературы Дубынин В. А. Регуляторные системы организма человека. М. : Дрофа, 2003. Дудьев В. П. Психомоторика: cловарь-справочник, 2008 г. Мэгун Г. Бодрствующий мозг. М. : Мир, 1965. Росси Я. А. , Цанкетти А. Ретикулярная формация ствола мозга. М. : ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М. : Изд-во Моск. ун-та, 1987.
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
по предмету: Физиологии высшей нервной деятельности
на тему: «Физиология сна и бодрствования»
Москва 2010
Введение
1. Теории сна
1.1 Восстановительная теория сна
1.2 Циркадианная теория сна
1.3 Гуморальная теория
1.4 Подкорковая и корковая теории сна
2. Фазы и стадии сна
3. Нейромеханизмы сна
4. Различные уровни бодрствования
5. Сон у животных
Заключение
Введение
Сон и бодрствование являются функциональными основными состояниями, в которых происходит жизнь человека. Эти функциональные состояния, хотя и противоположны, но тесно взаимосвязаны и рассматривать их следует в едином цикле «сон - бодрствование». Каждый вечер, когда мы засыпаем, наше сознание выключается на несколько часов. Мы перестаем воспринимать все происходящее вокруг. Здоровые люди воспринимают сон как обычное явление, и поэтому редко задумываются над его значением и природой. Но, когда сон нарушается, это причиняет нам много неприятностей.
В последнее время интерес к проблеме сна значительно возрос. В наше быстротечное время с его информационными перегрузками и экологическими влияниями значительно возросло количество людей, страдающих бессонницей. Сколько и нужно ли вообще спать человеку? Чем вызван сон, какова его роль в организме? Эти и другие вопросы стали предметом изучения физиологии сна. Еще в XVI столетии известный врач Парацельс придерживался того мнения, что естественный сон должен продолжаться 8 часов.
Сон (somnus) - функциональное состояние мозга и всего организма человека и животных, имеющее отличные от бодрствования специфические качественные особенности деятельности центральной нервной системы и соматической сферы, характеризующиеся торможением активного взаимодействия организма с окружающей средой и неполным прекращением (у человека) сознаваемой психической деятельности.
Важнейшими признаками бодрствования являются сознание, мышление и двигательная активность. В течение каждых суток сон и бодрствование сменяют друг друга, образуя генетически детерминированный суточный цикл сна-бодрствования.
1. Теории сна
1.1 Восстановительная теория сна
Восстановительная теория исторически была связана с изучением депривации сна и последствиями данного явления. Результатами депривации сна являются снижение работоспособности, ухудшается настроение, повышаются пороги чувствительности к сенсорным стимулам.
Все эти симптомы снимаются в случае здорового полноценного сна - в этом заключается восстановительная функция сна.
Также во время сна увеличивается секреция гормона роста, активируются анаболические процессы и происходит репаративное восстановление белковых молекул клеток.
Один из вариантов данной теории был разработан Павловым, считавшим, что сон по своей сути является распространяющимся в коре больших полушарий процесс охранительного торможения.
Однако эта теория была опровергнута впоследствии исследованиями, в ходе которых происходила регистрации электрической активности нейронов и показавшими, что их активность во сне не меньше, чем при бодрствовании.
Также она не подтверждается при сопоставлении продолжительности сна у разных видов млекопитающих с их физической активностью и скоростью обменных процессов.
1.2 Циркадианная теория сна
В контексте данной теории цикл сна и бодрствования рассматривается как результат управления циркадного ритма с помощью эндогенного механизма, не зависящий от внешних обстоятельств и определяемый как внутренние биологические часы.
Циркадный ритм - это связанный с естественным чередованием дня и ночи 24-часовой ритм.
Большинство имеющихся фактов указывает на то, что главным координатором биоритмологических процессов является гипоталамус. Водителем циркадных ритмов являются супрахиазмальные ядра (СХЯ) гипоталамуса, расположенные над зрительным перекрестом.
Они являются одними из двоих первичных синхронизаторов биологических ритмов, инициируя возникновение медленноволнового сна, регулируя интенсивность секреции гормона роста и скорость выделения кальция из организма.
Другой из синхронизаторов представлен в одной из областей вентромедиальных ядер (ВМЯ) гипоталамуса и служит в качестве регулятора быстроволнового сна, интенсивности секреции кортикостероидов, температуры тела и выделения калия из организма.
На данный момент две эти теории принято считать не как противоречащие теории, а как взаимодополняющие.
1.3 Гуморальная теория
В качестве причины сна данная теория рассматривает вещества, появляющиеся в крови при длительном бодрствовании.
Доказательством этой теории служит эксперимент, при котором бодрствующей собаке переливали кровь животного, лишенного сна в течение суток. Животное-реципиент немедленно засыпало.
В настоящее время удалось идентифицировать некоторые гипногенные вещества, например пептид, вызывающий дельта-сон. Но гуморальные факторы не могут рассматриваться как абсолютная причина возникновения сна. Об этом свидетельствуют наблюдения за поведением двух пар неразделившихся близнецов.
У них разделение нервной системы произошло полностью, а системы кровообращения имели множество анастомозов. Эти близнецы могли спать в разное время: одна девочка, например, могла спать, а другая бодрствовала.
1.4 Под корковая и корковая теории сна
При различных опухолевых или инфекционных поражениях подкорковых, особенно стволовых, образований мозга, у больных отмечаются различные нарушения сна - от бессонницы до длительного летаргического сна, что указывает на наличие подкорковых центров сна.
При раздражении задних структур субталамуса и гипоталамуса животные засыпали, а после прекращения раздражения они просыпались, что указывает на наличие в этих структурах центров сна.
Между лимбико-гипоталамическими и ретикулярными структурами мозга имеются реципрокные отношения. При возбуждении лимбико-гипоталамических структур мозга наблюдается торможение структур ретикулярной формации ствола мозга и наоборот.
При бодрствовании за счет потоков афферентации от органов чувств активируются структуры ретикулярной формации, которые оказывают восходящее активирующее влияние на кору больших полушарий. При этом нейроны лобных отделов коры оказывают нисходящие тормозные влияния на центры сна заднего гипоталамуса, что устраняет блокирующие влияния гипо-таламических центров сна на ретикулярную формацию среднего мозга. При уменьшении потока сенсорной информации снижаются восходящие активирующие влияния ретикулярной формации на кору мозга.
В результате чего устраняются тормозные влияния лобной коры на нейроны центра сна заднего гипоталамуса, которые начинают еще активнее тормозить ретикулярную формацию ствола мозга. В условиях блокады всех восходящих активирующих влияний подкорковых образований на кору мозга наблюдается медленноволновая стадия сна.
Гипоталамические центры за счет связей с лимбическими структурами мозга могут оказывать восходящие активирующие влияния на кору мозга при отсутствии влияний ретикулярной формации ствола мозга.
Эти механизмы составляют корково-подкорковую теорию сна (П.К.Анохин), которая позволила объяснить все виды сна и его расстройства. Она исходит из того, что состояние сна связано с важнейшим механизмом - снижением восходящих активирующих влияний ретикулярной формации на кору мозга.
Сон бескорковых животных и новорожденных детей объясняется слабой выраженностью нисходящих влияний лобной коры на гипоталамические центры сна, которые при этих условиях находятся в активном состоянии и оказывают тормозное действие на нейроны ретикулярной формации ствола мозга.
2. Фазы и с т адии сна
Наиболее распространенной и признанной теорией стадий сна признанна теория по Дементу и Клейтману, различающая их по изменениям глубины и частоты волн.
Выделяют две фазы сна - медленного (ФМС) и быстрого сна (ФБС); Иногда фазу быстрого сна называют парадоксальным сном. Эти названия обусловлены особенностями ритмики электроэнцефалограммы (ЭЭГ) во время сна - медленной активностью в ФМС и более быстрой в ФБС.
ФМС разделяется на 4 стадии, отличающиеся биоэлектрическими (электроэнцефалографическими) характеристиками и порогами пробуждения, являющимися объективными показателями глубины сна.
Первая стадия (дремота) характеризуется отсутствием на ЭЭГ б-ритма, являющегося характерным признаком бодрствования здорового человека, со снижением амплитуды и появлением низкоамплитудной медленной активности с частотой 3-7 в 1сек. (и - и д-ритмы). Могут регистрироваться ритмы и с более высокой частотой. На электроокулограмме возникают изменения биопотенциала, отражающие медленные движения глаз.
Вторая стадия (сон средней глубины) характеризуется ритмом «сонных веретен» с частотой 13-16 в 1 сек., то есть отдельные колебания биопотенциалов группируются в пачки, напоминающие форму веретена. В этой же стадии из фоновой активности четко выделяются 2 - 3-фазных высокоамплитудных потенциала, носящих название К-комплексов, нередко связанных с «сонными веретенами». К-комплексы регистрируются затем во всех стадиях ФМС. Амплитуда фоновой ритмики ЭЭГ при этом растет, а частота ее уменьшается по сравнению с первой стадией.
Для третьей стадии характерно появление на ЭЭГ медленной ритмики в д-диапазоне (то есть с частотой до 2 в 1 сек. и амплитудой 50-75 мкв и выше). При этом продолжают достаточно часто возникать «сонные веретена». Четвертая стадия (поведенчески наиболее глубокий сон) характеризуется доминированием на ЭЭГ высокоамплитудного медленного д-ритма.
Третья и четвертая стадии ФМС составляют так называемый дельта-сон.
ФБС отличается низкоамплитудной ритмикой ЭЭГ, а по частотному диапазону наличием как медленных, так и более высокочастотных ритмов (альфа - и бета-ритмов).
Характерными признаками этой фазы сна являются и так называемые пилообразные разряды с частотой 4-6 в 1 сек., быстрые движения глаз на электроокулограмме, в связи с чем эту фазу часто называют сном с быстрым движением глаз, а также резкое снижение амплитуды электромиограммы или полное падение тонуса мышц диафрагмы рта и шейных мышц.
3 . Нейром еханизмы сна
Одним из непроясненных вопросов на данный момент является вопрос о центрах сна. Несмотря на интенсивное изучение этого вопроса, точного ответа до сих пор нет.
Во второй половине нашего столетия прямое изучение нейронов, вовлеченных в регуляцию сна-бодрствования, показало, что нормальная работа таламо-кортикальной системы мозга, обеспечивающая сознательную деятельность человека в бодрствовании, возможна только при участии определенных подкорковых, так называемых активирующих, структур .
Благодаря их действиям в бодрствовании мембрана большинства кортикальных нейронов деполяризована на 10-15 мВ по сравнению с потенциалом покоя - (65-70) мВ. Только в состоянии этой тонической деполяризации нейроны способны обрабатывать информацию и отвечать на сигналы, приходящие к ним от других нервных клеток (рецепторных и внутримозговых).
Таких систем тонической деполяризации, или активации мозга, условных “центров бодрствования”, несколько - вероятно, пять или шесть. Они располагаются в различных частях мозга, а именно на всех уровнях мозговой оси: в ретикулярной формации ствола, в области голубого пятна и дорзальных ядер шва, в заднем гипоталамусе и базальных ядрах переднего мозга. Нейроны этих отделов выделяют медиаторы - глутаминовую и аспарагиновую кислоты, ацетилхолин, норадреналин, серотонин и гистамин, активность которых регулируют многочисленные пептиды, находящиеся с ними в одних и тех же везикулах. У человека нарушение деятельности любой из этих систем не компенсируется за счет других, несовместимо с сознанием и приводит к коме.
В связи с этим логично было бы предположить, что при допущении существования цетров бодрствования, должны существовать и центры сна. Однако в последние годы выяснилось, что в сами “центры бодрствования” встроен механизм положительной обратной связи. Это особые нейроны, которые осуществляют торможение активирующих нейронов и сами тормозятся ими. Такие нейроны разбросаны по разным отделам мозга, хотя больше всего их в ретикулярной части черного вещества. Все они выделяют один и тот же медиатор - гамма-аминомасляную кислоту, главное тормозное вещество мозга. Стоит только активирующим нейронам ослабить свою деятельность, как включаются тормозные нейроны и ослабляют ее еще сильнее. В течение некоторого времени процесс развивается по нисходящей, пока не срабатывает некий “триггер” и вся система переключается либо в состояние бодрствования, либо парадоксального сна. Объективно этот процесс отражает смена картин электрической активности головного мозга (ЭЭГ) по ходу одного полного цикла сна человека (90 мин).
Все чаще в последнее время внимание ученых привлекает еще одна эволюционно древняя тормозная система головного мозга, использующей в качестве медиатора нуклеозид аденозин.
Японский физиолог О. Хаяйси с коллегами показали, что синтезируемый в мозге простагландин D2 участвует в модуляции аденозинэргических нейронов. Поскольку главный фермент этой системы - простагландиназа-D - локализован в мозговых оболочках и хороидном плексусе, очевидна роль этих структур в формировании определенных видов патологии сна: гиперсомнии при некоторых черепно-мозговых травмах и воспалительных процессах менингеальных оболочек, африканской “сонной болезни”, вызываемой трипаносомой, которая передается через укусы мухи цеце и пр. Если с точки зрения нейронной активности бодрствование - это состояние тонической деполяризации, то медленный сон - тоническая гиперполяризация. При этом направление движения через клеточную мембрану основных ионных потоков (катионов Na+, K+, Ca2+, анионов Cl-), а также важнейших макромолекул меняется на противоположное. Это приводит к выводу, что во время медленного сна восстанавливается мозговой гомеостаз, нарушенный в ходе многочасового бодрствования.
С этой точки зрения бодрствование и медленный сон - как бы “две стороны одной медали”. Периоды тонической деполяризации и гиперполяризации должны периодически сменять друг друга, чтобы сохранить постоянство внутренней среды головного мозга и обеспечить нормальную работу таламо-кортикальной системы - субстрата высших психических функций человека.
Отсюда ясно, почему в мозге нет единого “центра медленного сна” - это значительно уменьшило бы надежность всей системы, сделало бы ее более жестко детерминированной, полностью зависящей от “капризов” этого центра в случае каких-либо нарушений его работы. В некотором смысле, данный факт подтверждает восстановительную теорию сна.
При этом складывается совершенно другая картина в отношении парадоксального сна, который, в отличие от медленного сна, имеет ярко выраженную активную природу . Парадоксальный сон запускается из четко очерченного центра, расположенного в задней части мозга, в области варолиева моста и продолговатого мозга, а медиаторами служат ацетилхолин, глутаминовая и аспарагиновая кислоты. Во время парадоксального сна клетки мозга чрезвычайно активны, но информация от органов чувств к ним не поступает и не подается на мышечную систему. В этом и заключается парадоксальность этого состояния . Фрагменты полиграммы на разных стадиях показывают, что для смены стадий медленного сна характерно постепенное увеличение амплитуды и снижение частоты волн ЭЭГ, смена быстрых движений глаз медленными, вплоть до полного исчезновения (ЭОГ регистрируется на фоне ЭЭГ и выделены цветом), прогрессивное уменьшение амплитуды ЭМГ. При парадоксальном сне ЭЭГ такая же, как при бодрствовании, ЭОГ демонстрирует быстрые движения глаз, а ЭМГ почти не регистрируется.
В этом случае допустить, что при этом интенсивно перерабатывается информация, полученная в предшествующем бодрствовании и хранящаяся в памяти. Согласно гипотезе Жуве, в парадоксальном сне, пока непонятно как, в нейрологическую память передается наследственная, генетическая информация, имеющая отношение к организации целостного поведения. Подтверждением таких психических процессов служит появление в парадоксальном сне эмоционально окрашенных сновидений у человека, а также обнаруженный Жуве с сотрудниками и детально исследованный Э.Моррисоном с коллегами феномен демонстрации сновидений у подопытных кошек .
Они выяснили, что в мозге кошек имеется особая область, ответственная за мышечный паралич во время парадоксального сна. Если ее разрушить, подопытные кошки начинают показывать свой сон: убегать от воображаемой собаки, ловить воображаемую мышь и т.д. Интересно, что “эротические” сны у кошек никогда не наблюдались, даже в брачный сезон.
Хотя в парадоксальном сне некоторые нейроны ретикулярной формации ствола и таламо-кортикальной системы демонстрируют своеобразный рисунок активности, различия между мозговой деятельностью в бодрствовании и парадоксальном сне довольно долго выявить не удавалось. Это было сделано лишь в 80-е годы.
Оказалось, что из всех известных активирующих мозговых систем, которые включаются при пробуждении и действуют во время бодрствования, в парадоксальном сне активны лишь одна-две. Это системы, расположенные в ретикулярной формации ствола и базальных ядрах переднего мозга, использующие в качестве передатчиков ацетилхолин, глутаминовую и аспарагиновую кислоты. Все же остальные активирующие медиаторы (норадреналин, серотонин и гистамин) в парадоксальном сне не работают. Это молчание моноаминоэргических нейронов ствола мозга определяет различие между бодрствованием и парадоксальным сном, или на психическом уровне - различие между восприятием внешнего мира и сновидений .
4 . Различные уровни бодрствования
Отличительным свойством сознания после пробуждения и во время активной деятельности является быстрота реагирования, способность сфокусировать внимание на тех или иных, мобилизовать ресурсы памяти.
В тоже время при низкой активности сознание отсутствует, как в прочем и в случае чрезмерной активности. Поэтому наиболее продуктивным уровнем активности является оптимальный, а не высокий.
Для активного бодрствования характерна следующая особенность: концентрируя свое внимание на объекте, наиболее значимом для него на данный момент, он теряет способность к восприятию остальных объектов.
Избирательность внимания, направленного на отдельные объекты, выделяемых из общего фона, связана с ограниченным объема оперативной памяти., не способной вместить всю поступающую сенсорную информацию.
Но с появлением раздражителя, отвлекающего внимание человека, происходит переключение посредством механизма ориентировочного рефлекса, после чего при восприятии данного раздражителя происходит изменение электроэнцефалограммы в специфической сенсорной области коры, где характерный для пассивного бодрствования б-ритм сменяется в-ритмом - такая десинхронизация получила название б-ритма.
Избирательное внимание человека, которое направленно на один конкретный объект, проявляется активацией не только первичных, но и вторичных сенсорных и ассоциативных областей коры, что увеличивает наши ресурсы для изучения данного объекта.
5. Сон у животных
Любым животным, от самых примитивных до высших, сон необходим так же, как человеку.
Сон -- это не просто отдых, а особое состояние мозга, которое отражается в специфическом поведении животного. Спящее животное, во-первых, принимает характерную для вида сонную позу, во-вторых, его двигательная активность резко снижается, в-третьих, оно перестает реагировать на внешние раздражители, однако способно в ответ на внешнюю или внутреннюю стимуляцию проснуться.
Следуя этим внешним признакам сна, окажется, что спят очень многие животные, как высшие, так и низшие.
Жирафы спят на коленях, заворачивая шею вокруг ног; львы лежат на спине, сложив передние лапы на груди, крысы укладываются на бок, а хвостик закручивают к голове. Так же спят и лисы. Летучие мыши засыпают, только подвесившись вниз головой. Как спят кошки, видел любой человек -- на боку с вытянутыми лапками. Коровы спят стоя и с открытыми глазами. У дельфинов и китов два полушария мозга спят по очереди. А иначе водное млекопитающее может «проспать» вдох и задохнуться.
Столь же разнообразны и «сонные» привычки птиц. Но в отличие от млекопитающих у птиц сохраняется большая двигательная активность и мышечный тонус. Для того чтобы заснуть, птице не обязательно ложиться, она может спать и стоя, и сидя на яйцах. Кроме того, многие птицы спят на лету. Иначе во время трансокеанических перелетов и без того измученной птице пришлось бы еще и без сна обходиться. Мигрирующие птицы спят так: каждые 10-15 минут в середину стаи залетает одна из птиц и чуть-чуть шевелит крыльями. Ее несет воздушный поток, создаваемый всей стаей. Потом ее место занимает другая птица. Могут птицы спать не только на лету, но и «на плаву»: утки спят, не вылезая на берег. А попугаи спят, повиснув на ветке вниз головой.
Как выяснилось, спят не только теплокровные животные, но и холоднокровные -- ящерицы, черепахи, рыбы. Раньше считалось, что холоднокровные животные просто замирали с наступлением холодной ночи, а вовсе не спали. Действительно, температура окружающей среды снижается, вместе с ней снижается и температура тела животного, падает уровень метаболизма, животное становится вялым и, как следствие, засыпает. Оказалось, однако, что дело не только в снижении уровня метаболизма. При постоянной температуре рептилии тоже засыпают.
Спят не только теплокровные животные -- спят змеи и даже пчелы.
Засыпают и раки, и насекомые, причем их сон отвечает тем внешним критериям, которые определены для высших животных. Пять лет назад Джоан Хендрикс в Пенсильванском университете удалось снять на видео, как спят мушки дрозофилы. Оказалось, что ночью они засыпают на 4-5 часов, да еще и днем сиесту устраивают часа на полтора, а всего за сутки маленькие фруктовые мушки спят около 8 часов. При этом перед сном они расползаются каждая на свое отдельное место, отворачиваются головой от пищи, ложатся на брюшко и замирают. Только ножки подрагивают, и брюшко ритмично раздувается в такт дыханию. Чем не сон усталого человека?
Сон у животных, как показали многочисленные исследования последних лет, связан с так называемыми циркадными ритмами. В организме живого существа существуют специальные «биологические часы», но их циферблат обычно чуть больше или меньше 24 часов, это время и составляет циркадный цикл. Эти часы «заводятся» специальными фотозависимыми белками. Дневной свет активизирует светочувствительные рецепторы, возбуждение передается группе нейронов мозга с работающими часовыми генами. Часовые гены синтезируют специальные белки, и функция этих часовых белков -- тормозить работу часовых генов! Получается саморегуляторная обратная связь: чем больше синтезировано часовых белков, тем меньше работает часовых генов. И так до тех пор, пока работа часовых генов не остановится и синтез белков не прекратится. С течением времени эти белки разрушаются, и работа часовых генов возобновляется. Циркадный цикл настроен обычно на длину светового дня.
Любопытно, что часовые гены мухи дрозофилы и млекопитающих очень похожи. Это говорит о том, что циклы сна и бодрствования очень древнего происхождения. Но насколько они древние -- покажут только будущие генетические исследования циркадных циклов. Не исключено, что окажется, что и микробы спят. А пока что сенсацией стало открытие генов короткого сна у мух дрозофил и очень похожих на них генов короткого сна у людей. Гены короткого сна передаются по наследству, как свидетельствует английский сомнолог Джером Сигел. Обладатели этих генов имеют укороченный сон, всего по 4-5 часов, после которого они вполне жизнерадостны и дееспособны. Правда, мухи с мутацией короткого сна имели и укороченную жизнь -- умирали на 2-3 недели раньше своих нормально спящих товарищей. Возможно, что у короткоспящих людей -- та же печальная зависимость. Например, Наполеон, спавший очень мало, умер в 52 года. Вполне вероятно, что его ранняя смерть -- это результат не печали и депрессий от одиночества, а подпорченных часовых генов. Впрочем, на сегодняшний день это только гипотеза.
Заключение
Существует достаточно большое количество исследований по физиологии сна и бодрствования, что говорит о все более возрастающем интересе к данной проблематике вопроса. В связи с этим появляется большое количество различных теорий сна и бодрствования, таких как восстановительная, циркадианная, гуморальная теории. Этот список можно продолжать и дальше.
Выделяют две основные фазы сна - медленного и быстрого или парадоксального сна. В свою очередь их также можно разложить на отдельные стадии сна, которые отличаются различными физиологическими показателями.
Говоря об нейромеханизмах сна, то можно говорить о том, что бодрствование - это состояние тонической деполяризации, то медленный сон - тоническая гиперполяризация.
Это приводит к выводу, что во время медленного сна восстанавливается мозговой гомеостаз, нарушенный в ходе многочасового бодрствования. С этой точки зрения бодрствование и медленный сон - как бы “две стороны одной медали”. Периоды тонической деполяризации и гиперполяризации должны периодически сменять друг друга, чтобы сохранить постоянство внутренней среды головного мозга и обеспечить нормальную работу таламо-кортикальной системы - субстрата высших психических функций человека.
Состояние же бодрствования также можно подразделить на различные уровни активности в зависимости от физиологического состояния, в котором находится человек на момент регистрирования.
Так же большой интерес представляет сон животных. У различных животных в зависимости от различных показателей существуют разные сонные привычки. Также является достоверным то, что у животных суточные ритмы могут определятся так де как и у человека циркадными ритмами.
Подобные документы
Изучение особенностей бодрствования, как одного из нейрофизиологических процессов психических механизмов человека. Показатели ЭЭГ-исследований. Период бодрствования на разных возрастных этапах. Регуляция функциональных состояний на уровне целого мозга.
реферат , добавлен 18.06.2011
Четыре периода сна: фетишистский, теологический, метафизический и эмпирио-психолого-физиологический. История развития представления о системе сна и бодрствования. Синхронизирующий эффект. Циклическая организация фаз сна. Нейрохимические механизмы сна.
реферат , добавлен 06.11.2012
Физиология и фазы сна. Электрические колебания мозга на разных его стадиях и во время бодрствования. Сущность химической, кортикальной, ретикулярной, серотонинергической концепций и корково-подкорковой, энергетической и информационной теорий сна.
презентация , добавлен 25.10.2014
Сущность процессов роста и развития организма. Этапы и периоды онтогенеза. Физическое и психическое развитие человека на жизненном пути. Биологические ритмы, их показатели и классификация. Чередование сна и бодрствования как основной суточный цикл.
контрольная работа , добавлен 03.06.2009
Сущность, биологическое значение и основные функции сна. Учение о сне, разработанное И.П. Павловым. Влияние сна и его отсутствия на организм. Структура нормального сна у здорового человека. Изменение продолжительности сна и бодрствования с возрастом.
доклад , добавлен 07.06.2010
Теория функциональных систем и её значение в формировании условно-рефлекторных поведенческих реакций животных. Учение Павлова об условных рефлексах, процесс и механизм их образования. Строение и значение анализаторов. Основные системы организма.
лекция , добавлен 08.05.2009
В XX в. происходило обсуждение и осмысление теории Ч. Дарвина. Согласно теории, человек возник в результате естественного процесса эволюции живой природы, имеет животных предков и его потребности, естественно, возникли на основе потребностей животных.
реферат , добавлен 26.06.2008
Единство принципа строения и развития мира растений и мира животных. Первые этапы формирования и развития представлений о клетке. Основные положения клеточной теории. Школа Мюллера и работа Шванна. Развитие клеточной теории во второй половине XIX века.
презентация , добавлен 25.04.2013
Классификация различных регуляторных механизмов сердечно-сосудистой системы. Влияние автономной (вегетативной) нервной системы на сердце. Гуморальная регуляция сердца. Стимуляция адренорецепторов катехоламинами. Факторы, влияющие на тонус сосудов.
презентация , добавлен 08.01.2014
Теории образования временной связи условного рефлекса. Физиология кожной чувствительности человека. Стадии и механизм условного рефлекса. Афферентные раздражения кожно-кинестетического анализатора. Отношения между интенсивностью стимула и ответом.
Взаимодействием активирующих и инактивирующих структур осуществляется регуляция цикла «сон-бодрствование». Несмотря на то, что все высшие позвоночные животные спят, а человек проводит во сне не менее трети своей жизни, природа и назначение этого состояния оставались неизвестными на протяжении веков. Хорошо известна была лишь витальная необходимость сна. Научное исследование сна стало возможным лишь в ХХ веке, после появления методов исследования, позволяющих определять изменение физиологических, биохимических показателей у спящего человека и регистрировать электрическую активность мозга. Сон, как и бодрствование - это активный процесс, связанный с деятельностью особых сомногенных механизмов мозга. Функции сна многообразны – восстановление ресурсов, переработка информации, психологическая адаптация, экономия энергии и жизненных сил организма.
У человека чередование бодрствования и ночного сна – проявление циркадианных (околосуточных) биологических ритмов. Супрахиазматические ядра переднего гипоталамуса получают по коллатералям зрительных нервов информация об уровне освещенности и оказывают избирательные влияния на «центры сна» или «центры бодрствования». Эпифиз также участвует в регуляции их активности.
Основной «центр бодрствования» - ретикулярные ядра моста, в которые поступают сигналы от всех сенсорных систем; далее происходит оценка уровня «сенсорного давления» на ЦНС, и чем оно больше, тем уровень активности больше. Нейроны этих ядер, в том числе ацетилхолинэргические, имеют широкие связи с разными структурами ЦНС. Уровень бодрствования повышается при активации адренэргических нейронов голубого пятна. Наступление сна вызывается активностью серотонинэргических нейронов ядер шва и центрального серого вещества среднего мозга. Происходит реципрокное торможение центров бодрствования, а также снижение активности таламических глутаминэргических нейронов, что приводит к торможению КБП. На активность «центров сна» влияют изменения химического состава крови: появление некоторых токсинов, повышенная концентрация азотсодержащих «шлаков», а также колебания уровня глюкозы увеличивают возбуждение сомногенных ядер.
Электроэнцефалографические исследования показали, что естественный сон – это чередование фазы медленноволнового сна, во время которого сменяются 5 стадий, и парадоксального сна, или сна с быстрыми движениями глаз (БДГ, REM – rapid eyes movement). На каждой стадии на ЭЭГ регистрируются определенные ритмы и происходит углубление сна (рис.23).
Рис 23. Ритмы электроэнцефалограммы на различных стадиях сна
Для бодрствования характерны высокочастотные и низкоамплитудные ά – ритм (спокойное бодрствование, частота 8-12 Гц) и β-ритм (активное бодрствование, 15-30 Гц). На I стадии (A, стадия дремоты) на фоне ά – ритма появляются эпизоды q - ритма (частота 4-8 Гц). На II стадии (B, самый поверхностный сон) преобладает q-ритм, в конце появляются высокоамплитудные зубцы над прецентральной извилиной. III стадия (C, поверхностный сон) характеризуется появлением «сонных веретен» и К-комплексов. Реакция на слабые раздражители уже отсутствует, наблюдаются реакции «вздрагиваниия». На IV стадии (D, умеренно глубокий сон) отмечается высокоамплитудные δ-волны, но их частота еще довольно велика, до 3,5 Гц. На V стадии (E, глубокий сон) частота δ-ритма становится крайне низкой – 0,7-1,2 Гц, но появляются эпизодически мелкие ά–волны. На протяжении медленного сна снижается тонус мышц, артериальное давление, температура тела, урежается пульс и дыхание. На фоне самого глубокого сна на ЭЭГ появляется ритм, характерный для бодрствования, что говорит о наступлении парадоксального сна. Признак этой фазы - быстрые движения глаз (rapid eyes movement): комплексы из 5-50 движений с частотой 60-70 в минуту. Их появление связывают с возбуждением стволовых глазодвигательных ядер при активации «центров бодрствования» - гигантоклеточных ретикулярных ядер моста и голубого пятна. Во время парадоксального сна на фоне максимального расслабления мускулатуры отмечаются мышечные подергивания, повышение температуры тела, колебания вегетативных показателей. При заболеваниях желудочно-кишечного тракта, сердечно-сосудистой системы в эти периоды сна могут возникать боли и ухудшение состояния. После фазы с БДГ обычно следует переход к II стадии или пробуждение.
Полный цикл сна занимает 60-90 минут, на сон с БДГ приходится около 20 %. За ночь проходит 4-6 циклов, с каждым последующим циклом продолжительность БДГ- сна увеличивается. У детей до 3-х лет сон с БДГ составляет до 50%.
Чередование фаз сна - это тоже проявление биоритмов (ультрадианные ритмы). Колебания активности с периодом около 90 минут отмечается и во время бодрствования. По всей вероятности, фазы сна выполняют различные функции: медленноволновой сон в большей степени восстановительную, сон с БДГ – информационную и адаптационную. Развитый БДГ – сон есть только у млекопитающих. Его депривация приводит к нарушениям самочувствия и эмоционального состояния.
Тот факт, что наиболее употребляемыми медикаментами в мире после болеутоляющих являются снотворные и тонизирующие средства, говорит о том, что человечеству крайне необходимо иметь средства воздействия на систему сна и бодрствования.
При этом следует отметить, что все существующие на рынке снотворные и стимуляторы имеют массу побочных действий и противопоказаний и, что самое важное, далеко не всегда эффективны. Именно этот факт и является движущей силой в раскрытии механизмов регуляции сна для создания более эффективных и безопасных снотворных и тонизирующих лекарственных препаратов.

История исследования регуляции сна не такая долгая. Впервые серьезно задумались над причинами возникновения сна только в начале XIX века, когда французский философ и врач (1757 - 1808) предположил, что сон возникает в результате застоя крови в мозгу вследствие горизонтального положения тела.
Так родилась гемодинамическая (циркуляторная) теория сна.

Эти взгляды получили продолжение в конце XIX века, когда (1846 - 1910) изобрел специальную кровать-весы. Человека укладывали на эту кровать и при его засыпании заметили, что головной конец кровати приподнимался.

Это расценивалось как отток крови от головы во время сна, вследствие чего голова становилась легче. Не все согласились с результатами Моссо: многие физиологи утверждали обратное, что сон возникает в результате прилива крови к голове и приводили свои доводы.
Хотя все перечисленные взгляды и были ошибочными, но они задали правильное направление, а именно они связывали возникновение сна с кровообращением в головном мозге.

Первыми по-настоящему научными опытами по исследованию регуляции сна были эксперименты на собаках французского психолога (1881 - 1964) , одного из классиков сомнологии.
Суть эксперимента в том, что собак привязывали коротким поводком к стене, что не позволяло животным заснуть. На 10й день (а на 11й обычно наступала смерть) часть собак умерщвляли, тогда как другим давали выспаться и только потом умерщвляли.
Клетки мозга первой части собак оказались в крайне ужасном состоянии: наблюдалось жировое перерождение нервных центров, кровеносные сосуды кишели лейкоцитами, а у тех, кому давали выспаться изменений в нейронах не было вообще.
Был сделан вывод о том, что на клетки мозга действует некое переносимое кровью эндогенное вещество, вызывающее сон - гипнотоксин (сонный яд) . Для подтверждения своей гипотезы Пьерон провел другой эксперимент, в котором бралась кровь, спинномозговая жидкость и экстракт из головного мозга от спящей собаки и вводилась собаке бодрствующей - та немедленно засыпала.
Так родилась химическая (гуморальная) теория сна , которая в несколько видоизмененном виде существует до сих пор. И хотя Пьерону так и не удалось выделить гипнотоксин, уже ни у кого не вызывало сомнений его существование. Казалось поколебать химическую теорию сна невозможно. Однако были описаны случаи, к которым эту теорию невозможно было применить.

В частности, академик (1898-1974) наблюдал в 1940х годах за сросшимися девочками-близнецами, у которых одна на двоих кровеносная система, но разные головы и обнаружил, что девочки могут спать в разное время - факт, который рушил всю теорию.

Чтобы проверить эти наблюдения, швейцарский нейробиолог (1907 — 1996) в 1965 году создал на собаках модель сиамских близнецов. У двух собак было налажено перекрестное кровообращение: кровь от мозга одной собаки текла в туловище другой и наоборот.
Когда определенный участок мозга собаки раздражали и провоцировали ее засыпание, засыпала и другая собака. Монье даже удалось в 1974 году выделить из крови спящих кроликов неуловимый гипнотоксин - полипептид, введение которого животным неизменно вызывало сон, названный впоследствии пептид дельта сна (DSIP, Delta sleep- inducing peptide) .
За последние десятилетия помимо пептида дельта-сна обнаружены и другие гипногенные вещества, такие как аденозин, фактор Папенхаймера, серотонин и другие.
Было постулировано, что сон запускается химическими агентами, которые накапливаются в течение бодрствования в организме и, достигая критической точки, переключают состояние бодрствования на состояние сна.
Очевидно, что химическая теория верна, но как же быть с настоящими, а не модельными сиамскими близнецами? Разрешить это явное противоречие удалось только с позиций центральных механизмов регуляции сна, впервые высказанной и яро пропагандируемой Иваном Петровичем Павловым (1849 - 1936) в его вертикальной теории сна.

По этой теории развитие сна связано с развивающимся утомлением нейронов, что создает условия для возникновения процессов торможения. Согласно Павлову, сон - это разлитое генерализованное торможение коры головного мозга и подкорковых структур.
При этом центральные нейронные механизмы являются определяющими, а гуморальные - второстепенными. Все дело в том, что одна голова сиамских близнецов просто не хотела спать вследствие очагов возбуждения в коре головного мозга, препятствующих иррадиации торможения.
Это может иметь место, когда существуют стимулы, которые в данный момент времени кажутся более значимыми, чем сон (принцип доминанты Ухтомского) . За интересной книгой мы можем провести всю ночь не сомкнув глаз, хотя концентрация гипногенных веществ в крови уже достаточно высока, чтобы запустить процесс засыпания.
По современным представлениям регуляция сна - это сложный нейрогуморальный процесс, имеющий локализованный анатомический субстрат. Среди факторов, вызывающих сон выделяют:
- Эндогенные факторы - вещества, накапливающиеся за период бодрствования, так называемые гипногенные вещества (серотонин, норадреналин, аденозин, дельта-пептид и т.д.) ;
- Циркадианные факторы, регулируемые биологическими часами - вещества с околосуточным ритмом ;
- Условнорефлекторные факторы (привычка ложиться спать в определенное время и т.д.) ;
- Безусловнорефлекторные факторы (темнота, покой, определенное положение тела, окружающая температура, атмосферное давление, монотонные стимулы на сенсорные системы - чтение книги, прослушивание музыки и т.д.) .
Эндогенные факторы сна

Одним из мощных эндогенных регуляторов сна является открытый в 1958 году профессором (1920 — 2007) главный регулятор суточных ритмов - .
Он вырабатывается эпифизом в ночное время. Причем и у дневных, и у ночных, и у сумеречных животных мелатонин вырабатывается только в темноте, то есть в ночное время и блокируется на свету. Его выработка целиком находится под контролем супрахиазматического ядра (СХЯ) гипоталамуса, небольшого скопления нервных клеток, представляющих собой наши «биологические часы».
Отношения между СХЯ и эпифизом реципрокные, а мелатонин начинает вырабатываться эпифизом примерно за час до отхода ко сну при неярком освещении. По современным представлениям считается, что мелатонин напрямую не регулирует сон, скорее он создает некую предрасположенность к отходу ко сну, реализуя седативный эффект. Поэтому прием мелатонина в качестве снотворного неэффективен.

Выработка мелатонина в течение суток.